You Cannot Out-Chemistry Your Energy Stack
In the first piece of this series, we mapped human energy as a five-tier system: starting at the rotary motor of ATP synthase deep inside your mitochondria, moving up through the neurochemical dashboard, the hormonal blueprint, the macroscopic inputs of light and food, and finally reaching the psychological apex of purpose.
Under normal conditions, that system regulates itself with remarkable precision. It produces, distributes, and consumes energy without you having to think about any of it.
The problem is that humans have never been particularly good at leaving a well-regulated system alone.
For as long as we've existed, we've introduced external molecules to rewrite the brain's commands. The morning espresso. The 4pm nootropic. The evening glass of wine. The weekend combination of THC, alcohol, and a few other things that adds up to a 9am Monday wreck. The pharmaceutical that fixes one symptom and creates another. The crystal meth pipe that promises three days of feeling unstoppable and delivers thirty days of a black hole instead.
Every one of those compounds, without a single exception, does the same fundamental thing. It rewrites the Level 2 dashboard. It tells the brain to spend ATP it wouldn't otherwise have spent, or to refuse spending ATP it would otherwise have released.
This piece is the mechanism itself: exactly how each major class of compound bends the stack, what it actually does to the dopamine, acetylcholine, GABA, and adenosine system, and why the biological bill always, always arrives.
1. The Neurochemical Command Center: How Brain Chemicals Rule ATP
To understand what these substances actually do, you have to look past muscle tissue and into the synaptic cleft, the microscopic gap between neurons where your perception of vitality actually gets decided.

Four molecules dominate the energy dashboard. Learn these, and the rest of this piece is just variations on a theme:
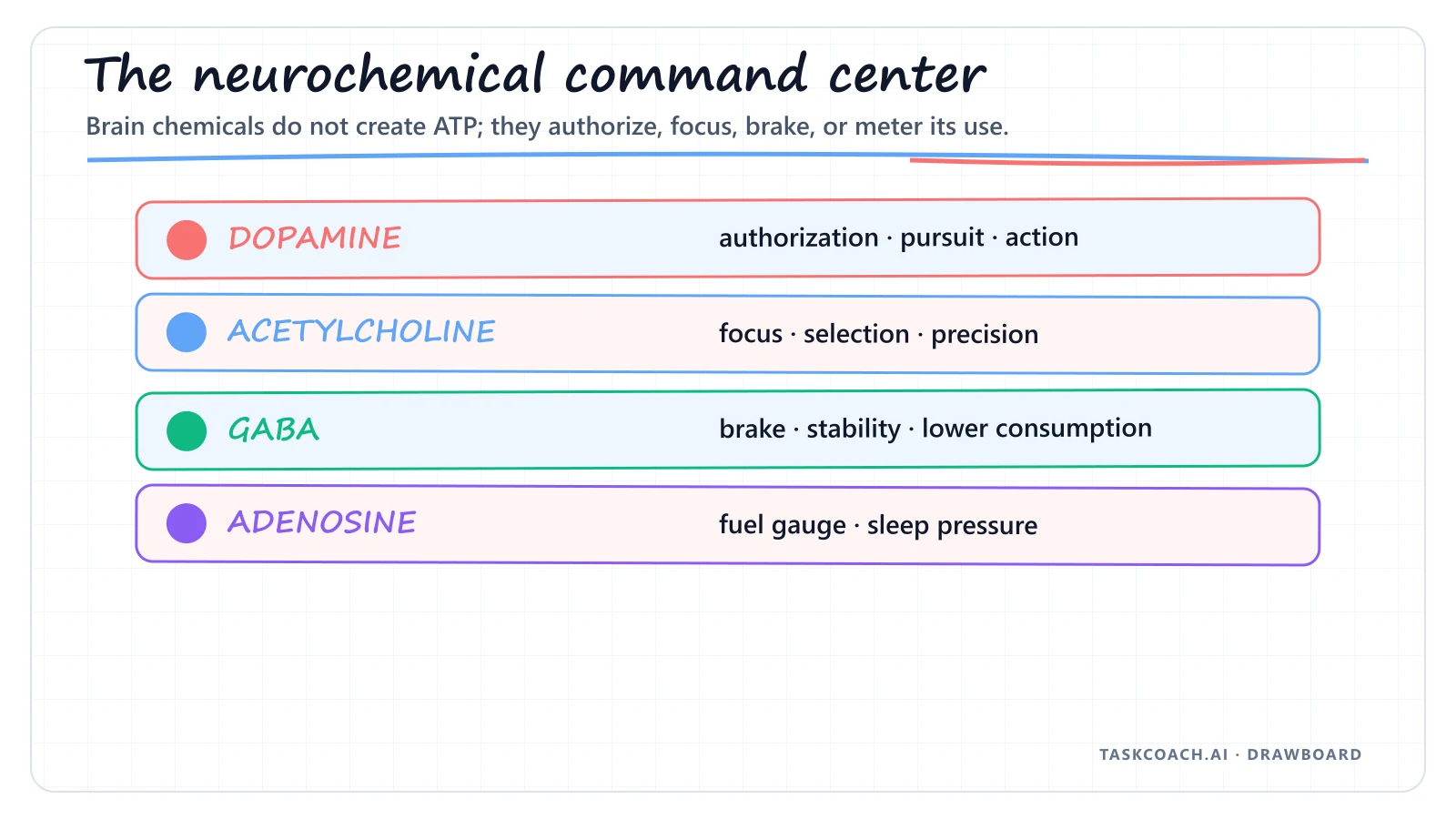
Dopamine: The Authorization Code
The most common misunderstanding in pop neuroscience is labeling dopamine the "pleasure molecule." Dopamine is really the molecule of anticipation, motivation, and pursuit. In energy terms, dopamine is the authorization code that unlocks ATP.

When dopamine spikes in the reward pathways, it tells the prefrontal cortex and the musculoskeletal system: a high-value reward is within reach, deploy resources now. Mitochondria ramp up cellular respiration, and glycogen and fat get converted into rapid-fire ATP.
When dopamine runs low, the vault stays locked. Cells drowning in glucose, mitochondria in perfect health, and the body simply refuses to spend. This is the structural reason chronic low motivation feels physical, not just mental.
Acetylcholine: The Lens of Computational Efficiency
If dopamine is the engine, acetylcholine is the lens. It governs attention, spatial awareness, and the precise focus of cognitive processing.
In energy terms, acetylcholine raises the signal-to-noise ratio of your brain. While you're thinking, only the neural circuits actually required for the task consume ATP, and everything else stays quiet. That razor-sharp feeling, being able to think hard for hours without exhaustion, is acetylcholine working at high efficiency.
GABA: The Systemic Brake
Gamma-aminobutyric acid, or GABA, is the brain's primary inhibitory neurotransmitter. Its job is to slow down neural activity by letting negatively charged chloride ions into the neuron, which makes it harder for the cell to fire.
GABA matters precisely because the brain is such a massive consumer of energy, using roughly 20% of your resting ATP. Without GABA forcing the system into a lower gear, neurons would fire continuously and asynchronously, leading to cellular exhaustion, oxidative stress, and excitotoxicity. GABA protects your energy reserves by forcing the machine to rest.
Adenosine: The Cellular Metronome
As your cells burn ATP throughout the day, a structural byproduct builds up in the brain: adenosine. The longer you've been awake, the more of it has accumulated.
Adenosine acts as your body's internal sleep-pressure gauge. It binds to A₁ and A₂A receptors and systematically dampens the release of dopamine and norepinephrine, essentially whispering that the currency has been spent, it's time to enter low-power standby and flush the metabolic waste.
This last molecule is where most of the manipulation begins.
2. The Day-to-Day Modulators: Tilting the Cellular Micro-Climate
Caffeine: Blinding the Energy Gauge
Caffeine is a master structural imposter. Its shape is almost identical to adenosine.

When you drink coffee, caffeine enters the synaptic cleft and parks directly inside your adenosine receptors as a competitive antagonist. The brake goes silent.
Caffeine does not generate a single molecule of ATP. It leaves your actual energy reserves completely untouched. It just mutes the cellular alarm that tells you you're running low. With the adenosine signal blocked, your existing dopamine and norepinephrine flow uninhibited, and a mild stress response releases some cortisol.
You feel alert. But it's an energetic illusion. You've taken out a high-interest loan against your future vitality. When the caffeine eventually clears, the backlog of adenosine that kept accumulating the whole time slams into the receptors all at once. That's your 3pm crash.
For a combination protocol that works around this elegantly, see the L-theanine and caffeine deep-work stack.
Synthetic Nootropics: Optimizing the Machinery
Modern nootropics like phenylpiracetam don't force a stressful neurotransmitter dump. Instead, they work as Positive Allosteric Modulators (PAMs) of AMPA and NMDA glutamate receptors. Glutamate is your brain's primary excitatory accelerator.
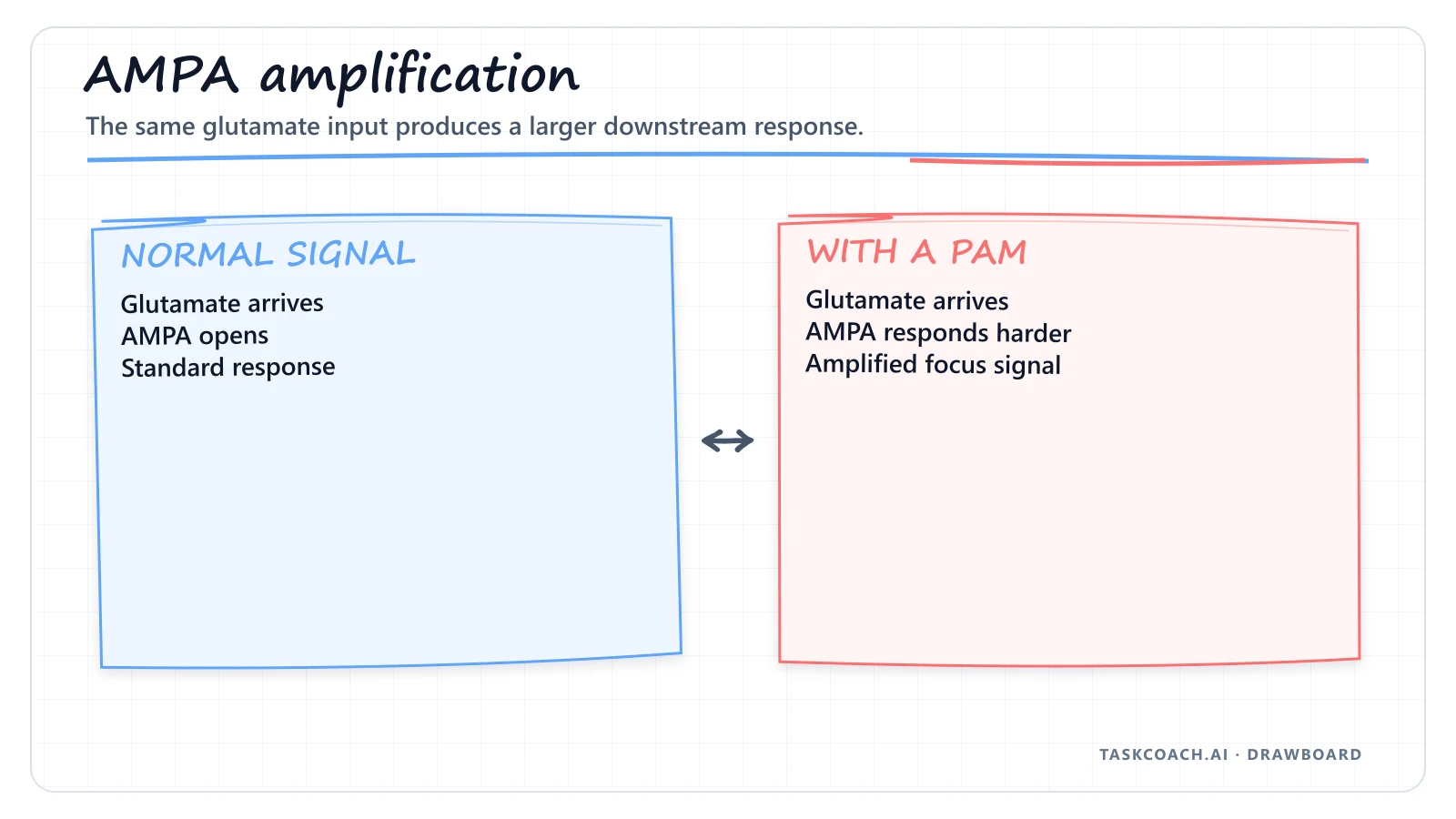
By changing the physical shape of these receptors, PAMs make them hyper-sensitive to the glutamate already floating around in your brain. At the same time, phenylpiracetam increases the density of acetylcholine receptors in the hippocampus.
By sensitizing both pathways at once, you get faster brainwaves and sharper focus without demanding emergency ATP reserves. You're optimizing the computational efficiency of the machine you already have. One caveat: tolerance and downregulation still apply here, so treat this as a tool, not a daily habit.
3. The Downward Arc: Depressants and the Amotivational Drift
To appreciate how powerful the neurochemical dashboard really is, look at the molecules that systematically dismantle it.
Alcohol: A Three-Phase Demolition
Alcohol (ethanol) is a profound CNS depressant that warps the brain's balance of excitation and inhibition in a very specific arc.
Phase 1: The disinhibited rush (the first twenty minutes or so). Alcohol triggers a transient release of dopamine and beta-endorphins. The frontal lobe's filters drop, behavioral inhibition relaxes, and the brain misreads the lack of friction as a surplus of expressive energy. This is the social-glow phase.
Phase 2: The GABA slam. As blood alcohol rises, ethanol binds heavily to GABA-A receptors, massively amplifying their inhibitory signal. At the same time, it plugs into NMDA glutamate receptors, blocking the brain's primary accelerator. Brakes locked, gas disconnected. Coordination fails. Speech slurs. ATP consumption drops, but only because the system is being forced into a chemical coma.
Phase 3: Mitochondrial suffocation (the next morning). As the liver breaks down ethanol, it produces acetaldehyde, a highly toxic byproduct that directly attacks the protein complexes inside the mitochondrial electron transport chain. Your cellular power plants effectively suffocate. Baseline ATP production plummets.
Meanwhile, your brain, panicking after hours of chemical suppression, releases a massive wave of stored glutamate: the glutamate rebound. This is the classic hangover state, hypersensitivity to light and sound, racing thoughts ("hangxiety"), physical incapacitation because your cells are starved of ATP while your brain is simultaneously over-amped.
The Anti-Motivational Drift: Cannabis and Doomscrolling
There's a quieter form of dashboard manipulation that doesn't involve any single substance: chronic exposure to high-dopamine, low-effort inputs. Prolonged heavy cannabis (THC) use. Hours a day of short-form video. Endless porn. Junk food on demand. Each one delivers a small flood of dopamine for almost no effort at all.
The brain has a protective response to this. When it's flooded with cheap rewards, it ruthlessly cuts back its own dopamine receptors, a process called downregulation.
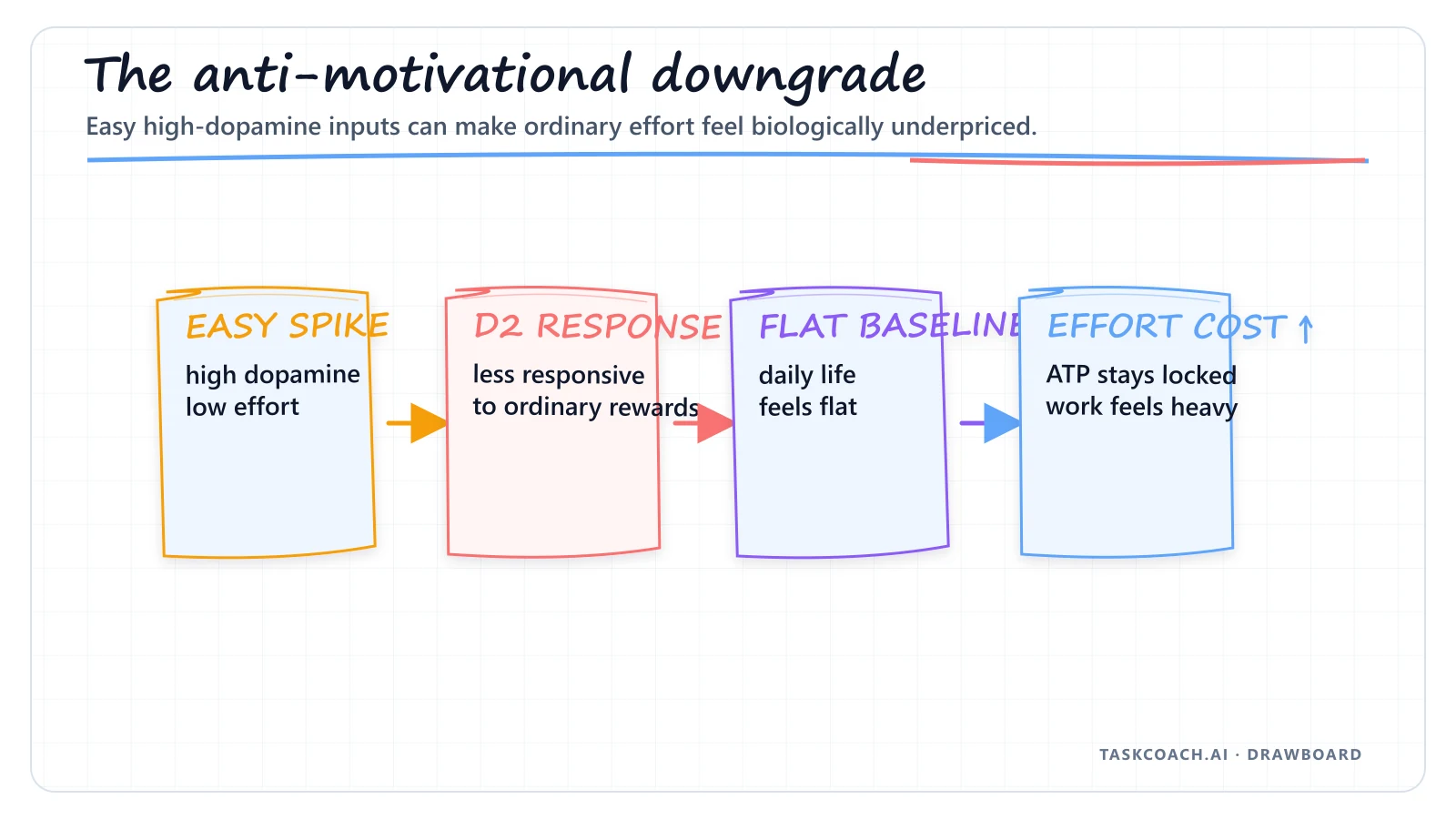
The result is a flattened dopamine baseline. When it's time to exercise, ship the project, or train the skill, the brain looks at the effort cost, looks at its own downregulated reward signal, and simply refuses to issue the authorization code.
The person experiences this as a heavy, immovable wall of apathy. Mitochondria are perfectly capable of producing ATP. Muscles are intact. The neurochemical command to actually ignite that energy just never arrives.
This is the structural reason a 30-day dopamine reset works: receptor density partially restores itself. For the supplement-supported version, see the NAC and Mucuna dopamine reboot stack. For the behavioral protocol, see the dopamine detox piece.
4. The Nuclear Options: Cocaine and Crystal Meth
When you cross into high-potency stimulants, you're no longer tweaking the dashboard. You're ripping it out of the wall.
Cocaine: The Synaptic Traffic Jam
Under normal conditions, once dopamine has delivered its motivational message across the synaptic cleft, a transporter protein called the Dopamine Transporter (DAT) acts like a microscopic vacuum cleaner, sucking the excess dopamine back into the cell to be recycled.
Cocaine binds directly to the DAT and jams the vacuum completely shut.
Every drop of dopamine you naturally release now has nowhere to go. It piles up in the synaptic cleft, slamming into the receiving receptors again and again. The result is an intense, ego-inflating rush of hyper-alertness, manic confidence, and a massive surge of perceived physical energy. Norepinephrine transporters get jammed the same way, so heart rate spikes, blood vessels constrict, and the whole physiology gets forced into maximum predatory survival mode.
You feel like a god, because your brain is registering a continuous, unmitigated reward signal, forcing Level 1 of the stack to empty its tanks. The next day, the receptors are exhausted. They were never built for that volume of signal. Tolerance comes fast. The crash comes faster.
Crystal Meth: The Total Intracellular Meltdown
If cocaine jams the vacuum cleaner, crystal meth (methamphetamine) turns the vacuum into a leaf blower running in reverse while setting the warehouse on fire.
Methamphetamine slips directly through cellular membranes; it doesn't need an open gate, since it's that lipid-soluble. Once inside the pre-synaptic neuron, it invades the VMAT-2 storage vesicles, the protective vaults where dopamine and norepinephrine are normally kept so they don't damage the cell's interior. Meth ruptures these vesicles, spilling massive reactive pools of neurotransmitter directly into the cytoplasm.
Then it gets worse. Meth forces the Dopamine Transporter to run completely in reverse. Instead of recycling dopamine back into the cell, the transporter starts pumping the massive internal flood out into the synaptic cleft, independent of any actual thought or environmental trigger.
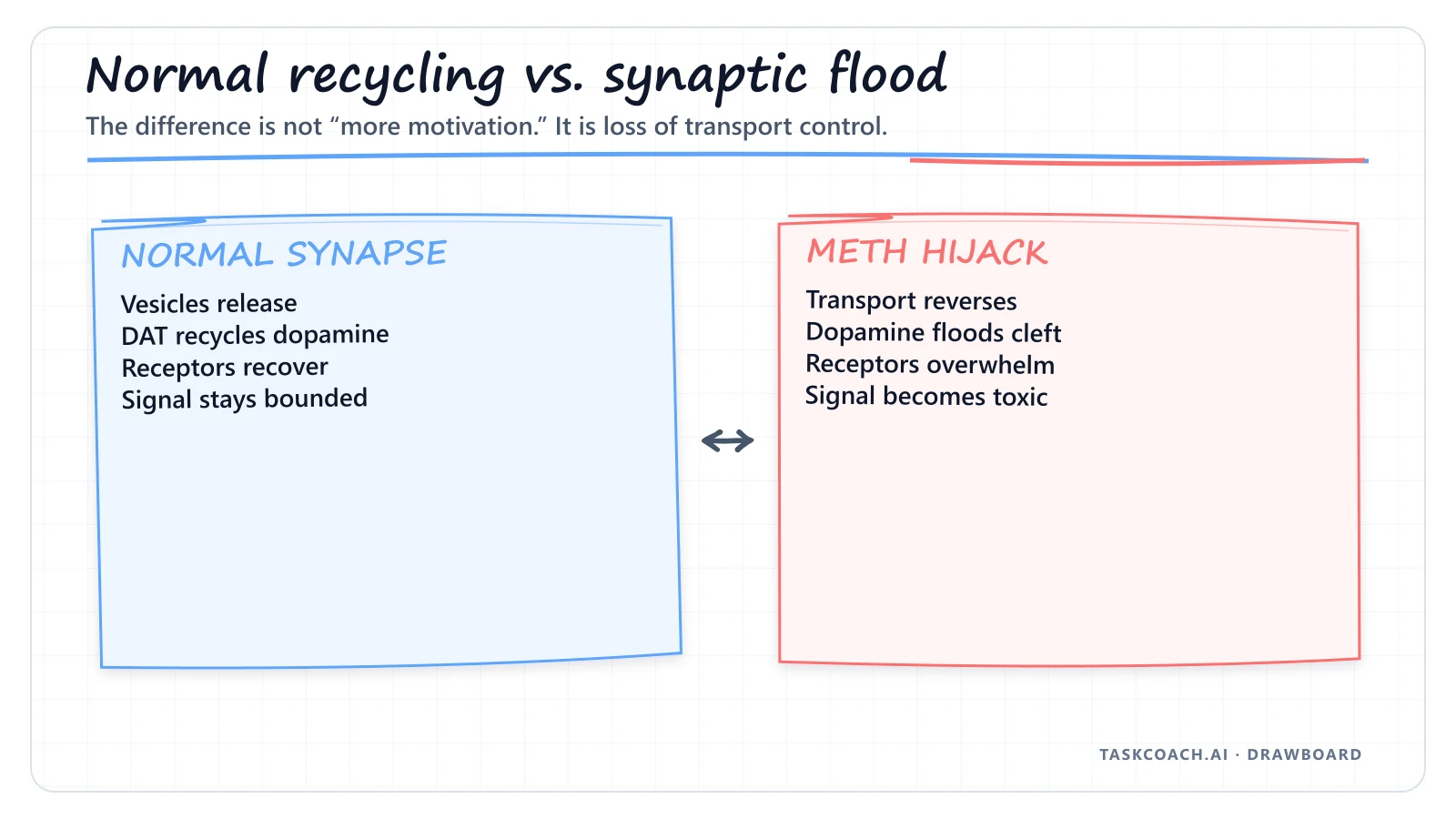
The synaptic dopamine concentration spikes by roughly 1200%, sustained for up to 24 hours. The neurochemical dashboard isn't modulated here. It's detonated.
The Catastrophic ATP Bankruptcy
Sustaining this artificial state of hyper-metabolism demands a toll that biological tissue simply cannot pay safely.
Because neurons are firing continuously at manic frequency, their delicate electrical gradients get wiped out. To keep them from dying, the sodium-potassium pumps (Na⁺/K⁺-ATPase), already the single largest consumer of ATP in the human body, have to run at maximum, around the clock, just to reset the cell's charge.
To fuel that, mitochondria get pushed into permanently hyper-accelerated cellular respiration. That extreme rate of production generates an overwhelming storm of reactive oxygen species (ROS), which are essentially cellular free radicals. These free radicals tear through the mitochondrial membranes and permanently destroy the protein complexes of the electron transport chain.
Your cellular power plants are literally burned out by their own hyper-production.
When the drug clears, the user enters an energetic black hole. The mitochondria have been structurally gutted by the free-radical storm. The body has lost its baseline capacity to generate ATP. Combined with severe dopamine receptor downregulation, the user experiences profound physical prostration, chronic fatigue, and anhedonia, a total biological inability to manufacture energy or feel motivated about anything.
The entire stack has been gutted.
The Immutable Law of Bio-Energetics
Whether you're optimizing the dashboard with a nootropic, blinding the gauge with caffeine, dampening the system with depressants, or watching the structural destruction caused by methamphetamine, the underlying law stays the same:
You cannot cheat the energy stack without paying the toll.
Substances that truly optimize human vitality do it by repairing the underlying machinery at the bottom of the stack: improving mitochondrial membrane integrity, reducing systemic inflammation, sensitizing receptors back toward baseline. They invest in the infrastructure. Their compounding interest works for you across years.
Substances that simulate energy do it by forcing the upper tiers to spend ATP the body hasn't safely accounted for. They pull the internal fire alarm to deplete tomorrow's reserves for a few hours of advantage today. Their compounding interest works against you.
True, sustainable, high-amplitude vitality has never been a product of chemical coercion. It's the natural byproduct of a well-maintained, biologically supported system, the one you read about in the first piece, which is what you're actually optimizing. The substances are sometimes useful, sometimes destructive, and almost never the place real change actually comes from.
The cup of coffee feels like the obvious move. The light exposure, the sleep architecture, the daily protocol that touches all five tiers, that's the actual move. The next piece in this series is the full end-to-end blueprint for that protocol: every supplement, every drill, every dose, start to finish.
This series is the underlying biology of what we're building at TaskCoach.AI. The supplements and biohacks people chase are downstream. The behavioral protocols that touch the bottom of the stack, light, sleep, dopamine hygiene, meaning, exercise, are what produce sustained vitality. A coaching system that holds those protocols across the seven life pillars, across months, is the difference between knowing about your energy stack and actually living inside an optimized one.